Der Hauptbestandteil der Zellwand. Zellenwand. Struktur. Funktionen. Zellwände von Algen
ZELLWAND, Zellmembran einzelliger, kolonialer und mehrzelliger Pilze und Pflanzen. In mehrzelligen Pflanzen bilden alle Zellwände und Interzellularräume eine einzige Gesamtorganismusstruktur (Apoplast), die den Stofftransport von Zelle zu Zelle gewährleistet. Die Dicke der Zellwand variiert zwischen verschiedenen Zellen (bei Pflanzen beispielsweise von 0,1 Mikrometer bis zu mehreren zehn Mikrometern). Die Stoffe, aus denen es besteht, werden hauptsächlich durch Polysaccharide repräsentiert. Sie bilden eine Matrix (amorphe Substanz) und darin enthaltene dünne Filamente unterschiedlicher Größe – Mikro- und Makrofibrillen, die praktisch nicht dehnbar sind und für die Zugfestigkeit der Zellwand sorgen. Solche Mikro- und Makrofibrillen bestehen aus parallelen linearen unverzweigten Molekülen von Chitin (in den meisten Pilzen), Cellulose (in einigen Pilzen und den meisten Pflanzen), Xylanen (in einer Reihe von Grün- und Rotalgen) und Mannanen (in einigen Grünalgen). Sie machen zwischen „4 % und „90 % der Trockenmasse der Zellwand aus, was von der taxonomischen Zugehörigkeit des Organismus, der Gewebeart und dem Alter der Zelle abhängt. Die Matrix basiert auf verzweigten und unverzweigten Molekülen aus Peptidoglykanen, Mannanen (Pilzen), Pektinen und Hemizellulosen (höhere Pflanzen), Alginaten und Fucoidan (Braunalgen), die durch verschiedene Bindungen zu einem durchbrochenen dreidimensionalen Netzwerk verbunden sind, das durchlässig ist zu wässrigen Lösungen. Die Matrix umfasst auch Proteine (einschließlich Glykoproteine), deren Rolle in Zellwänden kaum verstanden ist.
Die Bildung einer Zellwand beginnt mit der Ablagerung einer Matrixschicht auf der Oberfläche der Plasmamembran, deren Biopolymere im Inneren der Zelle synthetisiert und durch Exozytose an ihre Oberfläche freigesetzt werden. Die Synthese von Mikro- und Makrofibrillen erfolgt aus vom Protoplasten abgesonderten Monomervorläufern mithilfe mobiler Proteinkomplexe. Letztere dringen durch die Membran ein und sind mit den darunter liegenden Mikrotubuli des Zytoskeletts verbunden. Bei einzelligen und kolonialen Organismen ist die Anfangsschicht der Matrix frei von Mikro- und Makrofibrillen und oft verschleimt; in Sporen ist es mit Mineralien und/oder Polyphenolen verkrustet oder durch das sogenannte Sporopollenin ersetzt; Bei mehrzelligen Organismen wird die ursprüngliche Matrix zu einer Interzellularplatte. Anschließend bildet der Protoplast selbst die Zellwände, deren Matrix eine direkte Fortsetzung der ursprünglichen Schicht darstellt. Bei Pilzen bildet sich in der Dicke der Zellwand eine Mittelschicht aus Peptidoglykanen ohne Fibrillen. Bei Pflanzen gibt es primäre und sekundäre Zellwände. Der erste entsteht während des Zellwachstums und endet kurz danach; enthält eine kleine Anzahl zufällig ausgerichteter Mikro- und Makrofibrillen, die es der Wand ermöglichen, sich beim Zellwachstum zu dehnen. Zellen einiger Arten höherer Pflanzen bilden auf der primären Zellwand von innen eine sekundäre Zellwand mit eng, parallel zueinander angeordneten Mikro- und Makrofibrillen mit einer wohldefinierten Schichtstruktur. Die Fibrillen verschiedener Schichten der sekundären Zellwand sind in unterschiedliche Richtungen ausgerichtet, wodurch sie undehnbar wird. Zellwände können einer Verkalkung (z. B. bei „kalkhaltigen“ Grün- und Rotalgen), einer Verkieselung (z. B. bei Schachtelhalmen), einer Verholzung (bei Gefäßpflanzen), einer Suberisierung (bei höheren Pflanzen) und einem Ersatz der Pektin-Hemicellulose-Matrix unterliegen Kallose (in Siebfeldern Phloemelemente).
Die Zellwand übernimmt die Funktionen des äußeren Skeletts der Zelle, einer Schutzhülle, eines Regulators des Wasserhaushalts und ist an Stoffwechselprozessen und der Übertragung von Signalen aus der Umgebung in die Zelle beteiligt. Hemizellulosen der Zellwand im Endosperm von Samen sind eine Reservesubstanz für den Embryo (Kaki, Palme). Einige Zellen höherer Pflanzen haben im reifen Zustand keinen Protoplasten und werden ausschließlich durch eine Zellwand repräsentiert (z. B. Phellemzellen, leitende Xylemelemente); Holz besteht hauptsächlich aus der Zellwand abgestorbener Zellen. Tatsächlich sind es Zellwände, die als Rohstoffe für die Holz-, Zellstoff- und Papierindustrie, die Herstellung von Viskose sowie die Herstellung von Agar und Alginaten für die Medizin-, Lebensmittel- und Kosmetikindustrie dienen. Informationen zur Zellwand von Bakterien finden Sie in den Artikeln Bakterien, Gram-negative und Gram-positive Bakterien.
(Murein) und es gibt zwei Arten: grampositiv und gramnegativ. Die grampositive Zellwand besteht ausschließlich aus einer dicken Peptidoglycanschicht, die eng an der Zellmembran anliegt und von Teichon- und Lipoteichonsäuren durchdrungen ist. Beim gramnegativen Typ ist die Peptidoglycanschicht viel dünner; zwischen ihr und der Plasmamembran befindet sich ein periplasmatischer Raum, und außen ist die Zelle von einer anderen Membran umgeben, dargestellt durch die sogenannte. Lipopolysaccharid und ist ein pyrogenes Endotoxin gramnegativer Bakterien.
Zellwände von Pilzen
Pilzzellwände bestehen aus Chitin und Glucanen.
Zellwände von Algen
Die meisten Algen haben eine Zellwand aus Zellulose und verschiedenen Glykoproteinen. Einschlüsse zusätzlicher Polysaccharide sind von großer taxonomischer Bedeutung.
Zellwände höherer Pflanzen
Das wichtigste Unterscheidungsmerkmal einer Pflanzenzelle ist das Vorhandensein einer starken Zellwand, deren Hauptbestandteil Zellulose ist. Die Zellwand höherer Pflanzen ist eine komplexe, überwiegend polymere extrazelluläre Matrix, die jede Zelle umgibt. Eine Pflanzenzelle ohne Zellwand wird Protoplast genannt. In den Zellwänden von Pflanzen gibt es Vertiefungen – Poren, durch die zytoplasmatische Tubuli verlaufen – Plasmodesmen, die den Kontakt zwischen benachbarten Zellen und den Stoffaustausch zwischen ihnen durchführen.
Die chemische Zusammensetzung und die räumliche Organisation von Zellwandpolymeren unterscheiden sich zwischen verschiedenen Arten, Zellen verschiedener Gewebe derselben Pflanze und manchmal zwischen verschiedenen Teilen der Wand um denselben Protoplasten.
Darüber hinaus verändert sich die Struktur der Zellwand während der Ontogenese des pflanzlichen Organismus. Die primäre Zellwand wird während der Teilung gebildet und bleibt während des Zellwachstums erhalten. Die Bildung der sekundären Zellwand erfolgt an der Innenseite der Primärwand und ist mit dem Ende des Wachstums und der Spezialisierung (Differenzierung) der Pflanzenzellen verbunden. Außerhalb der primären Zellwand, zwischen den Primärwänden zweier benachbarter Zellen, befindet sich eine Mittelschicht (hauptsächlich bestehend aus Calcium- und Magnesiumsalzen von Pektinsubstanzen). Die primäre Zellwand höherer Pflanzen besteht aus drei interagierenden, aber strukturell unabhängigen dreidimensionalen Polymernetzwerken. Das Kernnetzwerk besteht aus Cellulosefibrillen und den Hemicellulosen (oder vernetzenden Glykanen), die sie verbinden. Das zweite Netzwerk besteht aus Pektinstoffen. Das dritte Netzwerk wird in der Regel durch Strukturproteine der Zellwand repräsentiert. Es ist auch zu beachten, dass bei Pflanzen der Commelinids-Gruppe (einer Gruppe in APG-Systemen) und bei Vertretern der Familie der Chenopodiaceae die primäre Zellwand eine erhebliche Menge an aromatischen Substanzen (Hydroxyzimtsäuren, hauptsächlich Ferulasäure) enthält P- kumarovaya). In diesem Fall sind Hydroxyzimtsäuren bei Vertretern der Commelinids-Gruppe an vernetzende Glykane (an Glucuronoarabinoxylane) und in der Familie der Chemopodiaceae an Pektinsubstanzen (an Rhamnogalacturonans I) gebunden.
Pflanzliche Zellwände erfüllen eine Reihe von Funktionen: Sie verleihen der Zelle Stabilität für strukturelle und mechanische Unterstützung, sie geben der Zelle ihre Form, die Richtung ihres Wachstums und letztendlich die Morphologie der gesamten Pflanze. Die Zellwand wirkt auch dem Turgor, also dem osmotischen Druck, entgegen, wenn zusätzliches Wasser in die Pflanze gelangt. Zellwände schützen vor dem Eindringen von Krankheitserregern aus der Umwelt und speichern Kohlenhydrate für die Pflanze.
siehe auch
|
||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||
Schreiben Sie eine Bewertung zum Artikel „Zellwand“
Auszug zur Charakterisierung der Zellwand
„Wie einfach, wie wenig Aufwand nötig ist, um so viel Gutes zu bewirken, dachte Pierre, und wie wenig uns das interessiert!“Er freute sich über die ihm entgegengebrachte Dankbarkeit, schämte sich jedoch, sie anzunehmen. Diese Dankbarkeit erinnerte ihn daran, wie viel mehr er für diese einfachen, freundlichen Menschen hätte tun können.
Der Chefdirektor, ein sehr dummer und listiger Mann, der den klugen und naiven Grafen vollkommen verstand und wie ein Spielzeug mit ihm spielte, sah, welche Wirkung die vorbereiteten Techniken auf Pierre hatten, und wandte sich entschiedener an ihn mit Argumenten über die Unmöglichkeit und, Am wichtigsten ist die Unnötigkeit der Befreiung der Bauern, die auch ohne sie völlig glücklich waren.
Pierre stimmte dem Manager insgeheim zu, dass man sich kaum glücklichere Menschen vorstellen könne und dass Gott weiß, was sie in der Wildnis erwartete; aber Pierre bestand, wenn auch widerstrebend, auf dem, was er für fair hielt. Der Verwalter versprach, seine ganze Kraft einzusetzen, um den Willen des Grafen auszuführen, wohlwissend, dass der Graf ihm niemals vertrauen würde, und zwar nicht nur hinsichtlich der Frage, ob alle Maßnahmen ergriffen worden seien, um Wälder und Ländereien zu verkaufen und vom Rat abzulösen , würde aber wahrscheinlich auch nie fragen oder erfahren, wie die gebauten Gebäude leer stehen und die Bauern weiterhin mit Arbeit und Geld alles geben, was sie von anderen geben, also alles, was sie geben können.
Als Pierre von seiner Reise in den Süden zurückkam, verwirklichte er im glücklichsten Gemütszustand seine langjährige Absicht, seinen Freund Bolkonsky aufzusuchen, den er seit zwei Jahren nicht gesehen hatte.
Bogucharovo lag in einer hässlichen, flachen Gegend, bedeckt mit Feldern und gefällten und unbeschnittenen Tannen- und Birkenwäldern. Der Hof des Herrenhauses lag am Ende einer geraden Linie, entlang der Hauptstraße des Dorfes, hinter einem neu gegrabenen, vollgefüllten Teich, dessen Ufer noch nicht mit Gras bewachsen waren, inmitten eines jungen Waldes, dazwischen standen mehrere große Kiefern.
Der Hof des Herrenhauses bestand aus einer Tenne, Nebengebäuden, Ställen, einem Badehaus, einem Nebengebäude und einem großen Steinhaus mit halbrundem Giebel, das sich noch im Bau befand. Rund um das Haus wurde ein junger Garten angelegt. Die Zäune und Tore waren stark und neu; unter dem Baldachin standen zwei Feuerrohre und ein grün gestrichenes Fass; Die Straßen waren gerade, die Brücken waren stark und mit Geländern versehen. Alles trug den Eindruck von Ordentlichkeit und Sparsamkeit. Als die Diener, die sich trafen, gefragt wurden, wo der Prinz wohnte, zeigten sie auf ein kleines, neues Nebengebäude, das ganz am Rande des Teiches stand. Anton, der alte Onkel von Prinz Andrei, ließ Pierre aus der Kutsche steigen, sagte, der Prinz sei zu Hause und führte ihn in einen sauberen, kleinen Flur.
Pierre war beeindruckt von der Bescheidenheit des kleinen, wenn auch sauberen Hauses nach den hervorragenden Bedingungen, in denen er seinen Freund das letzte Mal in St. Petersburg gesehen hatte. Er betrat eilig die immer noch nach Kiefern duftende, unverputzte, kleine Halle und wollte weitergehen, doch Anton schlich auf Zehenspitzen nach vorne und klopfte an die Tür.
- Nun, was ist da? – war eine scharfe, unangenehme Stimme zu hören.
„Gast“, antwortete Anton.
„Bitten Sie mich zu warten“, und ich hörte, wie ein Stuhl zurückgeschoben wurde. Pierre ging schnell zur Tür und stand Prinz Andrei gegenüber, der stirnrunzelnd und gealtert auf ihn zukam. Pierre umarmte ihn, hob seine Brille, küsste ihn auf die Wangen und sah ihn genau an.
„Das habe ich nicht erwartet, ich bin sehr froh“, sagte Prinz Andrei. Pierre sagte nichts; Er sah seinen Freund überrascht an, ohne den Blick abzuwenden. Er war beeindruckt von der Veränderung, die bei Prinz Andrei stattgefunden hatte. Die Worte waren liebevoll, ein Lächeln lag auf den Lippen und im Gesicht von Prinz Andrei, aber sein Blick war stumpf und tot, dem Prinz Andrei trotz seines offensichtlichen Wunsches keinen freudigen und fröhlichen Glanz verleihen konnte. Es ist nicht so, dass sein Freund abgenommen hat, blass geworden ist und reifer geworden ist; Aber dieser Blick und die Falte auf seiner Stirn, die lange Konzentration auf eine Sache ausdrückten, erstaunten und entfremdeten Pierre, bis er sich daran gewöhnte.
Wenn man sich nach einer langen Trennung trifft, kann das Gespräch, wie es immer passiert, nicht lange aufhören; Sie fragten und antworteten kurz über Dinge, von denen sie selbst wussten, dass sie ausführlicher besprochen werden sollten. Schließlich begann sich das Gespräch nach und nach auf das zuvor bruchstückhaft Gesagte zu konzentrieren, auf Fragen zu seinem früheren Leben, zu Plänen für die Zukunft, zu Pierres Reisen, zu seinen Aktivitäten, zum Krieg usw. Diese Konzentration und Depression, die Pierre bemerkte Im Blick von Prinz Andrei drückte sich das Lächeln, mit dem er Pierre zuhörte, noch stärker aus, besonders wenn Pierre mit lebhafter Freude über die Vergangenheit oder die Zukunft sprach. Es war, als ob Prinz Andrei an dem, was er sagte, teilhaben wollte, aber nicht konnte. Pierre begann zu spüren, dass Begeisterung, Träume, Hoffnungen auf Glück und Güte vor Prinz Andrei nicht angemessen waren. Er schämte sich, all seine neuen, freimaurerischen Gedanken zum Ausdruck zu bringen, insbesondere diejenigen, die durch seine letzte Reise in ihm erneuert und erregt worden waren. Er hielt sich zurück, hatte Angst, naiv zu sein; Gleichzeitig wollte er seinem Freund unwiderstehlich schnell zeigen, dass er nun ein ganz anderer, besserer Pierre war als der, der in St. Petersburg war.
Die Zellmembran ist ein typischer Bestandteil einer Pflanzenzelle und ein Produkt der lebenswichtigen Aktivität des Protoplasten.
Funktionen:
1. Starke und starre Zellmembranen dienen als mechanische Unterstützung für Pflanzenorgane.
2. Die Membran begrenzt die Ausdehnung des Protoplasten durch die Vakuole und die Größe und Form der reifen Zelle ändert sich nicht mehr.
3. Im äußeren Gewebe schützen Zellmembranen tiefer liegende Zellen vor dem Austrocknen.
4. Verschiedene Stoffe und Wasser können sich entlang benachbarter Zellwände von Zelle zu Zelle bewegen (Weg durch den Apoplasten).
5. Sie beeinflussen Absorption, Transpiration und Sekretion.
Zellwände sind normalerweise farblos und lassen Sonnenlicht leicht durch. Die Wände benachbarter Zellen werden durch Pektin zusammengehalten Mittelplatte. Die mittlere Lamina ist eine einzelne Schicht, die zwei benachbarten Zellen gemeinsam ist. Es handelt sich um eine leicht veränderte Zellplatte, die während des Prozesses der Zytokinese entstanden ist. Die mittlere Platte ist weniger bewässert und kann Ligninmoleküle enthalten. Durch den intrazellulären Druck können die Winkel der Zellwände abgerundet werden und es bilden sich Interzellularräume zwischen benachbarten Zellen. Alle Pflanzenzellwände, die miteinander verbunden sind und an wassergefüllte Interzellularräume angrenzen, sorgen für die Existenz einer kontinuierlich bewässerten Umgebung, in der sich wasserlösliche Substanzen frei bewegen können.
Struktur und chemische Zusammensetzung.
Primäre Zellwand.
Zunächst erscheint außerhalb des Plasmalemmas Primärzelle Wand.
Verbindung: Zellulose, Hemizellulose, Pektin und Wasser.
Die primären Zellwände benachbarter Zellen sind durch eine Protopektin-Mittelschicht verbunden. In der Zellwand werden lineare, sehr lange (mehrere Mikrometer) Cellulosemoleküle, die aus Glucose bestehen, zu Bündeln gesammelt – Mizellen, die wiederum zu Mikrofibrillen zusammengefasst werden – den dünnsten (1,5...4 nm) Fasern unbestimmter Länge. und dann in Makrofibrillen. Zellulose bildet ein mehrdimensionales Gerüst, das in eine amorphe, stark wasserhaltige Matrix aus nichtzellulosehaltigen Kohlenhydraten eingebettet ist: Pektine, Hemizellulosen usw. Es ist Zellulose, die für die Festigkeit der Zellwand sorgt. Mikrofibrillen sind elastisch und haben eine ähnliche Zugfestigkeit wie Stahl. Matrixpolysaccharide bestimmen Wandeigenschaften wie hohe Wasserdurchlässigkeit, gelöste kleine Moleküle und Ionen sowie starke Quellung. Dank der Matrix können sich Wasser und Stoffe entlang der aneinander angrenzenden Wände von Zelle zu Zelle bewegen (der Weg durch den Apoplasten entlang des „Freiraums“). Einige Hemizellulosen können als Speicherstoffe in den Zellwänden von Samen eingelagert werden.
Wandwachstum.
Bei der Zellteilung entsteht lediglich die Zellplatte neu. Beide Tochterzellen legen darauf ihre eigenen Wände an, die hauptsächlich aus Hemizellulose bestehen. In diesem Fall kommt es auch an der Innenfläche der übrigen zur Mutterzelle gehörenden Wände zur Wandbildung. Die Zellplatte wandelt sich in eine Mittelplatte um; sie ist meist dünn und kaum zu unterscheiden. Nach der Teilung tritt die Zelle aufgrund der Wasseraufnahme durch die Zelle und dem Wachstum der zentralen Vakuole in eine Elongationsphase ein. Der Turgordruck dehnt die Wand, in die Cellulosemizellen und Matrixsubstanzen eingebettet sind. Diese Wachstumsmethode wird aufgerufen Invagination, Implementierung. Die Membranen sich teilender und wachsender Zellen werden als primär bezeichnet. Sie enthalten bis zu 90 % Wasser, die Trockenmasse wird von Matrixpolysacchariden dominiert: Bei Dikotyledonen sind Pektine und Hemicellulosen zu gleichen Teilen vorhanden, bei Monokotyledonen hauptsächlich Hemicellulosen; Der Zellulosegehalt darf 30 % nicht überschreiten. Die Dicke der Primärwand beträgt nicht mehr als 0,1...0,5 Mikrometer.
Wenn das Zellwachstum endet, kann die Zellwand weiter wachsen, allerdings mit zunehmender Dicke. Dieser Vorgang wird als sekundäre Eindickung bezeichnet. In diesem Fall wird eine sekundäre Zellwand auf der Innenfläche der primären Zellwand abgelagert. Dadurch kommt es zum Wachstum der sekundären Zellwand Apposition, das Aufbringen neuer Cellulosemizellen auf die innere Oberfläche der Zellwand. Somit sind die jüngsten Schichten der Zellwand dem Plasmalemma am nächsten.
Bei einigen Zelltypen (viele Fasern, Tracheiden, Gefäßsegmente) ist die Bildung einer Sekundärwand die Hauptfunktion des Protoplasten; nach Abschluss der sekundären Verdickung stirbt er ab. Dies ist jedoch nicht notwendig. Die Sekundärwand übernimmt hauptsächlich mechanische, tragende Funktionen. Seine Zusammensetzung enthält deutlich weniger Wasser und es überwiegen Zellulose-Mikrofibrillen (40...50 % der Trockenmasse). In den Sekundärwänden von Flachsfasern und Baumwollhaaren kann der Zellulosegehalt bis zu 95 % betragen.
Mechanismus des Zellwandaufbaus. Die Zellwand wird durch die Aktivität des Protoplasten gebildet. Dementsprechend gelangen Stoffe von innen, von der Protoplastenseite her, in die Wand. Baustoffe – Zellulosemoleküle aus Pektin, Lignin und anderen Stoffen – sammeln sich an und werden teilweise in den Tanks des Golgi-Apparats synthetisiert. Verpackt in Vesikeln des Golgi-Apparats werden sie zum Plasmalemma transportiert. Nach dem Platzen platzt die Blase und ihr Inhalt erscheint außerhalb des Plasmalemmas. Die Vesikelmembran stellt die Integrität des Plasmalemmas wieder her. Dank der enzymatischen Aktivität des Plasmalemmas werden Cellulosefibrillen zur Zellwand zusammengefügt. Die vom Plasmalemma gebildeten Fibrillen liegen von innen übereinander, ohne sich zu verflechten. Eine wichtige Rolle in ihrer Ausrichtung spielen Mikrotubuli, die sich unter dem Plasmalemma parallel zu den sich bildenden Fibrillen befinden.
2. Poren. Veränderungen der Zellwand.
Poren. Bei der Bildung der primären Zellwand werden darin dünnere Bereiche unterschieden, in denen Zellulosefibrillen lockerer liegen. Die Tubuli der endoplasmatischen Kette durchdringen hier die Zellwände und verbinden benachbarte Zellen. Diese Bereiche werden aufgerufen primäre Porenfelder und die durch sie verlaufenden Tubuli des endoplasmatischen Retikulums Plasmodesmen .
Das Dickenwachstum erfolgt an der Zellwand ungleichmäßig; kleine Abschnitte der primären Zellwand bleiben an den Stellen der primären Porenfelder (Porenkanäle) unverdickt. Die Porenkanäle zweier benachbarter Zellen liegen sich meist gegenüber und werden durch einen Porenverschlussfilm – zwei primäre Zellwände mit dazwischen liegender Interzellularsubstanz – getrennt. Der Film enthält submikroskopische Öffnungen, durch die Plasmodesmen gelangen. Auf diese Weise, Die Zeit besteht aus zwei Porenkanälen und einem schließenden Film zwischen ihnen.
Plasmodesmen dringen in die Verschlussfilme der Poren ein. Jede Zelle enthält mehrere Hundert bis Zehntausende Plasmodesmen. Plasmodesmen kommen nur in Pflanzenzellen vor, wo feste Zellwände vorhanden sind. Plasmodesmen werden aus ER-Tubuli gebildet, die in der Zellplatte zwischen zwei Tochterzellen verbleiben. Wenn das ER beider Zellen wiederhergestellt wird, werden sie durch Plasmodesmen verbunden.
Plasmodesmen passieren den Plasmodesmalkanal im Verschlussfilm der Pore. Das Plasmalemma, das den Kanal auskleidet, und das Hyaloplasma zwischen ihm und den Plasmodesmen gehen in die Plasmalemmas und Hyaloplasmen benachbarter Zellen über. Somit sind die Protoplasten benachbarter Zellen durch Plasmodesmata-Kanäle und Plasmodesmata miteinander verbunden. Sie befördern den interzellulären Transport von Ionen und Molekülen sowie Hormonen. Die durch Plasmodesmen verbundenen Protoplasten der Zellen der Pflanze bilden ein einziges Ganzes – den Symplasten. Der Transport von Stoffen durch Plasmodesmen wird im Gegensatz zum apoplastischen Transport entlang von Zellwänden und Interzellularräumen als symplastisch bezeichnet.
Während des Lebens einer Zelle kann es zu Veränderungen an der Zellulosezellwand kommen.
Zu Beginn der Entwicklung des Lebens auf der Erde wurden alle Zellformen durch Bakterien repräsentiert. Sie nahmen organische Substanzen, die im Urmeer gelöst waren, über die Körperoberfläche auf.
Im Laufe der Zeit haben sich einige Bakterien daran angepasst, aus anorganischen Substanzen organische Substanzen herzustellen. Dazu nutzten sie die Energie des Sonnenlichts. Es entstand das erste Ökosystem, in dem diese Organismen als Produzenten fungierten. Dadurch gelangte der von diesen Organismen freigesetzte Sauerstoff in die Erdatmosphäre. Mit seiner Hilfe können Sie aus derselben Nahrung viel mehr Energie gewinnen und die zusätzliche Energie nutzen, um die Struktur des Körpers zu komplizieren: den Körper in Teile zu teilen.
Eine der wichtigsten Errungenschaften des Lebens ist die Trennung von Zellkern und Zytoplasma. Der Zellkern enthält Erbinformationen. Eine spezielle Membran um den Kern ermöglichte den Schutz vor unbeabsichtigter Beschädigung. Bei Bedarf erhält das Zytoplasma vom Zellkern Befehle, die das Leben und die Entwicklung der Zelle steuern.
Organismen, bei denen der Zellkern vom Zytoplasma getrennt ist, haben das nukleare Superreich gebildet (dazu gehören Pflanzen, Pilze und Tiere).
So entstand und entwickelte sich die Zelle – die Grundlage der Organisation von Pflanzen und Tieren – im Laufe der biologischen Evolution.
Schon mit bloßem Auge, oder noch besser unter einer Lupe, kann man erkennen, dass das Fruchtfleisch einer reifen Wassermelone aus sehr kleinen Körnern bzw. Körnern besteht. Dies sind Zellen – die kleinsten „Bausteine“, aus denen der Körper aller lebenden Organismen, einschließlich Pflanzen, besteht.
Das Leben einer Pflanze erfolgt durch die gemeinsame Aktivität ihrer Zellen, wodurch ein einziges Ganzes entsteht. Bei der Mehrzelligkeit von Pflanzenteilen kommt es zu einer physiologischen Differenzierung ihrer Funktionen, einer Spezialisierung verschiedener Zellen je nach ihrer Lage im Pflanzenkörper.
Eine Pflanzenzelle unterscheidet sich von einer tierischen Zelle dadurch, dass sie eine dichte Membran hat, die den inneren Inhalt von allen Seiten bedeckt. Die Zelle ist nicht flach (wie es normalerweise dargestellt wird), sondern sieht höchstwahrscheinlich wie eine sehr kleine Blase aus, die mit schleimigem Inhalt gefüllt ist.
Struktur und Funktionen einer Pflanzenzelle
Betrachten wir eine Zelle als strukturelle und funktionelle Einheit eines Organismus. Die Außenseite der Zelle ist mit einer dichten Zellwand bedeckt, in der sich dünnere Abschnitte, sogenannte Poren, befinden. Darunter befindet sich ein sehr dünner Film – eine Membran, die den Zellinhalt – das Zytoplasma – bedeckt. Im Zytoplasma gibt es Hohlräume – mit Zellsaft gefüllte Vakuolen. In der Mitte der Zelle oder in der Nähe der Zellwand befindet sich ein dichter Körper – ein Zellkern mit einem Nukleolus. Der Kern ist durch die Kernhülle vom Zytoplasma getrennt. Kleine Körper, sogenannte Plastiden, sind im Zytoplasma verteilt.
Struktur einer Pflanzenzelle
Struktur und Funktionen pflanzlicher Zellorganellen
| Organoid | Zeichnung | Beschreibung | Funktion | Besonderheiten |
Zellwand oder Plasmamembran | Farblos, transparent und sehr langlebig | Leitet Substanzen in die Zelle hinein und aus ihr heraus. | Die Zellmembran ist semipermeabel |
|
Zytoplasma | Dickflüssige Substanz | Alle anderen Teile der Zelle befinden sich darin | Ist in ständiger Bewegung |
|
Zellkern (wichtiger Teil der Zelle) | Rund oder oval | Sorgt für die Übertragung erblicher Eigenschaften auf Tochterzellen während der Teilung | Zentraler Teil der Zelle |
|
Kugelförmige oder unregelmäßige Form | Beteiligt sich an der Proteinsynthese | |||
 | Ein durch eine Membran vom Zytoplasma getrenntes Reservoir. Enthält Zellsaft | Überschüssige Nährstoffe und Abfallprodukte, die die Zelle nicht benötigt, sammeln sich an. | Während die Zelle wächst, verschmelzen kleine Vakuolen zu einer großen (zentralen) Vakuole |
|
Plastiden | Chloroplasten | Sie nutzen die Lichtenergie der Sonne und erzeugen aus Anorganischem Organisches | Die Form von Scheiben, die durch eine Doppelmembran vom Zytoplasma abgegrenzt sind |
|
Chromoplasten | Entsteht durch die Ansammlung von Carotinoiden | Gelb, Orange oder Braun |
||
 | Leukoplasten | Farblose Plastiden | ||
Atomhülle | Besteht aus zwei Membranen (äußere und innere) mit Poren | Trennt den Zellkern vom Zytoplasma | Ermöglicht den Austausch zwischen Zellkern und Zytoplasma |
Der lebende Teil einer Zelle ist ein membrangebundenes, geordnetes, strukturiertes System aus Biopolymeren und inneren Membranstrukturen, die an einer Reihe von Stoffwechsel- und Energieprozessen beteiligt sind, die das gesamte System als Ganzes aufrechterhalten und reproduzieren.
Ein wichtiges Merkmal ist, dass die Zelle keine offenen Membranen mit freien Enden hat. Zellmembranen begrenzen Hohlräume oder Bereiche immer und verschließen sie nach allen Seiten.
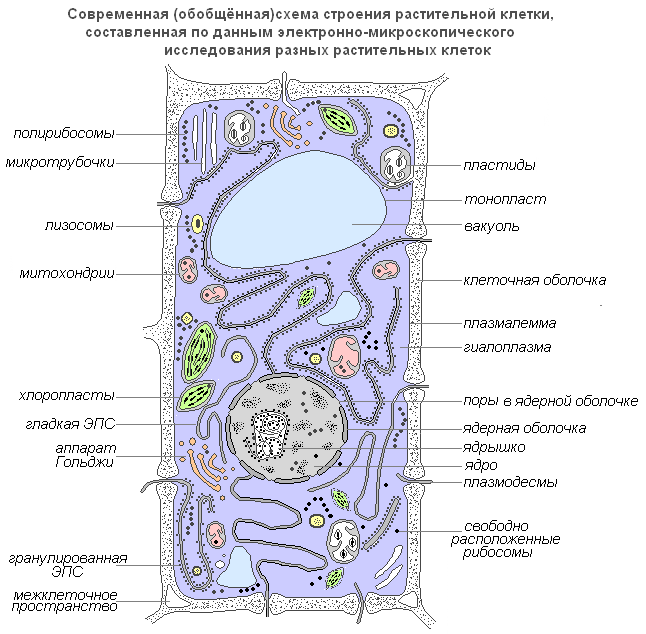
Modernes verallgemeinertes Diagramm einer Pflanzenzelle
Plasmalemma(äußere Zellmembran) ist ein ultramikroskopischer Film mit einer Dicke von 7,5 nm, der aus Proteinen, Phospholipiden und Wasser besteht. Dabei handelt es sich um einen sehr elastischen Film, der von Wasser gut benetzt wird und nach einer Beschädigung schnell seine Integrität wiederherstellt. Es hat eine universelle Struktur, d. h. typisch für alle biologischen Membranen. In Pflanzenzellen gibt es außerhalb der Zellmembran eine starke Zellwand, die für äußere Unterstützung sorgt und die Form der Zelle beibehält. Es besteht aus Ballaststoffen (Cellulose), einem wasserunlöslichen Polysaccharid.
Plasmodesmen Pflanzenzellen sind submikroskopisch kleine Röhrchen, die die Membranen durchdringen und mit einer Plasmamembran ausgekleidet sind, die so ohne Unterbrechung von einer Zelle zur anderen gelangt. Mit ihrer Hilfe kommt es zu einer interzellulären Zirkulation von Lösungen, die organische Nährstoffe enthalten. Sie übertragen auch Biopotentiale und andere Informationen.
Porami sogenannte Öffnungen in der Sekundärmembran, wo die Zellen nur durch die Primärmembran und die mittlere Lamina getrennt sind. Die Bereiche der Primärmembran und der Mittelplatte, die die benachbarten Poren benachbarter Zellen trennen, werden Porenmembran oder Verschlussfilm der Pore genannt. Der Verschlussfilm der Pore wird von plasmodesmalen Tubuli durchstoßen, in den Poren bildet sich jedoch meist kein Durchgangsloch. Poren erleichtern den Transport von Wasser und gelösten Stoffen von Zelle zu Zelle. Poren bilden sich in den Wänden benachbarter Zellen, meist einander gegenüber.
Zellmembran hat eine gut definierte, relativ dicke Schale aus Polysacchariden. Die Hülle einer Pflanzenzelle ist ein Produkt der Aktivität des Zytoplasmas. An seiner Entstehung sind der Golgi-Apparat und das endoplasmatische Retikulum aktiv beteiligt.

Struktur der Zellmembran
Die Basis des Zytoplasmas ist seine Matrix oder Hyaloplasma, ein komplexes farbloses, optisch transparentes kolloidales System, das zu reversiblen Übergängen vom Sol zum Gel fähig ist. Die wichtigste Aufgabe des Hyaloplasmas besteht darin, alle Zellstrukturen in einem einzigen System zu vereinen und die Interaktion zwischen ihnen in den Prozessen des Zellstoffwechsels sicherzustellen.
Hyaloplasma(oder zytoplasmatische Matrix) bildet die innere Umgebung der Zelle. Es besteht aus Wasser und verschiedenen Biopolymeren (Proteine, Nukleinsäuren, Polysaccharide, Lipide), von denen der Hauptteil aus Proteinen unterschiedlicher chemischer und funktioneller Spezifität besteht. Das Hyaloplasma enthält außerdem Aminosäuren, Monosaccharide, Nukleotide und andere niedermolekulare Substanzen.
Biopolymere bilden mit Wasser ein kolloidales Medium, das je nach Bedingungen sowohl im gesamten Zytoplasma als auch in seinen einzelnen Abschnitten dicht (in Form eines Gels) oder flüssiger (in Form eines Sols) sein kann. Im Hyaloplasma sind verschiedene Organellen und Einschlüsse lokalisiert und interagieren miteinander und mit der Hyaloplasmaumgebung. Darüber hinaus ist ihr Standort meist spezifisch für bestimmte Zelltypen. Über die Bilipidmembran interagiert das Hyaloplasma mit der extrazellulären Umgebung. Folglich ist Hyaloplasma eine dynamische Umgebung und spielt eine wichtige Rolle für die Funktion einzelner Organellen und das Leben von Zellen im Allgemeinen.
Zytoplasmatische Formationen - Organellen
Organellen (Organellen) sind strukturelle Bestandteile des Zytoplasmas. Sie haben eine bestimmte Form und Größe und sind obligatorische zytoplasmatische Strukturen der Zelle. Fehlen sie oder sind sie beschädigt, verliert die Zelle meist ihre Überlebensfähigkeit. Viele der Organellen sind zur Teilung und Selbstreproduktion fähig. Ihre Größe ist so klein, dass sie nur mit einem Elektronenmikroskop sichtbar sind.
Kern
Der Zellkern ist das prominenteste und meist größte Organell der Zelle. Es wurde erstmals 1831 von Robert Brown im Detail erforscht. Der Zellkern stellt die wichtigsten Stoffwechsel- und genetischen Funktionen der Zelle bereit. Die Form ist sehr unterschiedlich: Sie kann kugelförmig, oval, gelappt oder linsenförmig sein.
Der Zellkern spielt eine wichtige Rolle im Leben der Zelle. Eine Zelle, deren Kern entfernt wurde, scheidet keine Membran mehr aus und hört auf zu wachsen und Substanzen zu synthetisieren. Die Zerfalls- und Zerstörungsprodukte verstärken sich darin, wodurch es schnell stirbt. Die Bildung eines neuen Zellkerns aus dem Zytoplasma findet nicht statt. Neue Kerne entstehen nur durch Teilung oder Zerkleinerung des alten.
Der innere Inhalt des Kerns ist Karyolymphe (Kernsaft), der den Raum zwischen den Strukturen des Kerns ausfüllt. Es enthält einen oder mehrere Nukleolen sowie eine beträchtliche Anzahl von DNA-Molekülen, die mit bestimmten Proteinen – Histone – verbunden sind.

Kernstruktur
Nukleolus
Der Nukleolus enthält wie das Zytoplasma überwiegend RNA und spezifische Proteine. Seine wichtigste Funktion besteht darin, dass es Ribosomen bildet, die in der Zelle die Synthese von Proteinen durchführen.
Golgi-Apparat
Der Golgi-Apparat ist ein Organell, das in allen Arten eukaryontischer Zellen universell verbreitet ist. Es handelt sich um ein mehrschichtiges System flacher Membransäcke, die sich entlang der Peripherie verdicken und Blasenfortsätze bilden. Es befindet sich am häufigsten in der Nähe des Kerns.

Golgi-Apparat
Der Golgi-Apparat umfasst notwendigerweise ein System kleiner Vesikel (Vesikel), die von verdickten Zisternen (Scheiben) abgelöst werden und sich entlang der Peripherie dieser Struktur befinden. Diese Vesikel spielen die Rolle eines intrazellulären Transportsystems für spezifische Sektorgranula und können als Quelle für zelluläre Lysosomen dienen.
Zu den Funktionen des Golgi-Apparats gehört auch die Ansammlung, Trennung und Freisetzung von intrazellulären Syntheseprodukten, Abbauprodukten und toxischen Substanzen mithilfe von Vesikeln außerhalb der Zelle. Produkte der synthetischen Aktivität der Zelle sowie verschiedene Substanzen, die aus der Umgebung über die Kanäle des endoplasmatischen Retikulums in die Zelle gelangen, werden zum Golgi-Apparat transportiert, sammeln sich in diesem Organell an und gelangen dann in Form von Tröpfchen oder Körnern in das Zytoplasma und werden entweder von der Zelle selbst genutzt oder nach außen ausgeschieden. . In Pflanzenzellen enthält der Golgi-Apparat Enzyme für die Synthese von Polysacchariden und das Polysaccharidmaterial selbst, das zum Aufbau der Zellwand verwendet wird. Es wird angenommen, dass es an der Bildung von Vakuolen beteiligt ist. Der Golgi-Apparat wurde nach dem italienischen Wissenschaftler Camillo Golgi benannt, der ihn 1897 erstmals entdeckte.
Lysosomen
Lysosomen sind kleine, von einer Membran begrenzte Vesikel, deren Hauptfunktion darin besteht, die intrazelluläre Verdauung durchzuführen. Die Nutzung des lysosomalen Apparats erfolgt während der Keimung eines Pflanzensamens (Hydrolyse von Reservenährstoffen).

Struktur eines Lysosoms
Mikrotubuli
Mikrotubuli sind membranöse, supramolekulare Strukturen, die aus spiralförmig oder geraden Reihen angeordneten Proteinkügelchen bestehen. Mikrotubuli erfüllen überwiegend eine mechanische (motorische) Funktion und sorgen für die Beweglichkeit und Kontraktilität der Zellorganellen. Sie befinden sich im Zytoplasma, geben der Zelle eine bestimmte Form und sorgen für die Stabilität der räumlichen Anordnung der Organellen. Mikrotubuli erleichtern die Bewegung von Organellen an Orte, die durch die physiologischen Bedürfnisse der Zelle bestimmt werden. Eine beträchtliche Anzahl dieser Strukturen befindet sich im Plasmalemma, in der Nähe der Zellmembran, wo sie an der Bildung und Ausrichtung von Cellulose-Mikrofibrillen pflanzlicher Zellwände beteiligt sind.

Mikrotubuli-Struktur
Vakuole
Die Vakuole ist der wichtigste Bestandteil pflanzlicher Zellen. Es ist eine Art Hohlraum (Reservoir) in der Masse des Zytoplasmas, gefüllt mit einer wässrigen Lösung aus Mineralsalzen, Aminosäuren, organischen Säuren, Pigmenten, Kohlenhydraten und vom Zytoplasma durch eine Vakuolenmembran – den Tonoplasten – getrennt.
Nur in den jüngsten Pflanzenzellen füllt Zytoplasma den gesamten inneren Hohlraum aus. Während die Zelle wächst, verändert sich die räumliche Anordnung der zunächst zusammenhängenden Zytoplasmamasse erheblich: Es entstehen kleine, mit Zellsaft gefüllte Vakuolen und die gesamte Masse wird schwammig. Bei weiterem Zellwachstum verschmelzen einzelne Vakuolen und drängen die Zytoplasmaschichten an die Peripherie, wodurch die gebildete Zelle meist eine große Vakuole enthält und sich das Zytoplasma mit allen Organellen in der Nähe der Membran befindet.
Wasserlösliche organische und mineralische Verbindungen der Vakuolen bestimmen die entsprechenden osmotischen Eigenschaften lebender Zellen. Diese Lösung einer bestimmten Konzentration ist eine Art osmotische Pumpe für das kontrollierte Eindringen in die Zelle und die Freisetzung von Wasser, Ionen und Metabolitenmolekülen aus dieser.
In Kombination mit der Zytoplasmaschicht und ihren Membranen, die sich durch semipermeable Eigenschaften auszeichnen, bildet die Vakuole ein wirksames osmotisches System. Osmotisch bestimmt werden Indikatoren lebender Pflanzenzellen wie osmotisches Potenzial, Saugkraft und Turgordruck.

Struktur der Vakuole
Plastiden
Plastiden sind die größten (nach dem Zellkern) zytoplasmatischen Organellen, die nur den Zellen pflanzlicher Organismen innewohnen. Sie kommen nicht nur in Pilzen vor. Plastiden spielen eine wichtige Rolle im Stoffwechsel. Sie sind durch eine doppelte Membranhülle vom Zytoplasma getrennt und einige Arten verfügen über ein gut entwickeltes und geordnetes System innerer Membranen. Alle Plastiden sind vom gleichen Ursprung.
Chloroplasten- die häufigsten und funktionell wichtigsten Plastiden photoautotropher Organismen, die photosynthetische Prozesse durchführen, die letztendlich zur Bildung organischer Substanzen und der Freisetzung von freiem Sauerstoff führen. Chloroplasten höherer Pflanzen haben eine komplexe innere Struktur.

Chloroplastenstruktur
Die Größe der Chloroplasten in verschiedenen Pflanzen ist nicht gleich, aber im Durchschnitt beträgt ihr Durchmesser 4–6 Mikrometer. Chloroplasten können sich unter dem Einfluss der Bewegung des Zytoplasmas bewegen. Darüber hinaus wird unter dem Einfluss von Licht eine aktive Bewegung von Chloroplasten vom Amöbentyp in Richtung der Lichtquelle beobachtet.
Chlorophyll ist der Hauptstoff der Chloroplasten. Dank Chlorophyll können grüne Pflanzen Lichtenergie nutzen.
Leukoplasten(farblose Plastiden) sind klar definierte zytoplasmatische Körper. Ihre Größe ist etwas kleiner als die Größe von Chloroplasten. Ihre Form ist auch gleichmäßiger und nähert sich der Kugelform.
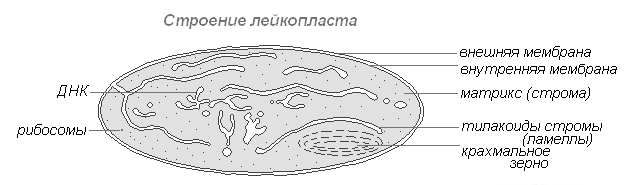
Leukoplast-Struktur
Kommt in Epidermiszellen, Knollen und Rhizomen vor. Bei Beleuchtung verwandeln sie sich sehr schnell in Chloroplasten mit entsprechender Veränderung der inneren Struktur. Leukoplasten enthalten Enzyme, mit deren Hilfe aus überschüssiger Glukose, die bei der Photosynthese entsteht, Stärke synthetisiert wird, deren Großteil sich in Form von Stärkekörnern in Speichergeweben oder Organen (Knollen, Rhizome, Samen) ablagert. Bei manchen Pflanzen lagern sich Fette in Leukoplasten ab. Die Reservefunktion von Leukoplasten äußert sich gelegentlich in der Bildung von Reserveproteinen in Form von Kristallen oder amorphen Einschlüssen.
Chromoplasten In den meisten Fällen handelt es sich um Derivate von Chloroplasten, gelegentlich auch von Leukoplasten.

Chromoplast-Struktur
Die Reifung von Hagebutten, Paprika und Tomaten geht mit der Umwandlung von Chloro- oder Leukoplasten der Pulpazellen in Caratinoidplasten einher. Letztere enthalten überwiegend gelbe Plastidenpigmente – Carotinoide, die im reifen Zustand darin intensiv synthetisiert werden und farbige Lipidtröpfchen, feste Kügelchen oder Kristalle bilden. In diesem Fall wird Chlorophyll zerstört.
Mitochondrien
Mitochondrien sind für die meisten Pflanzenzellen charakteristische Organellen. Sie haben eine variable Form von Stäbchen, Körnern und Fäden. 1894 von R. Altman mit einem Lichtmikroskop entdeckt und die innere Struktur später mit einem Elektronenmikroskop untersucht.

Die Struktur der Mitochondrien
Mitochondrien haben eine Doppelmembranstruktur. Die äußere Membran ist glatt, die innere bildet Auswüchse unterschiedlicher Form – Röhren in Pflanzenzellen. Der Raum im Mitochondrium ist mit halbflüssigem Inhalt (Matrix) gefüllt, der Enzyme, Proteine, Lipide, Kalzium- und Magnesiumsalze, Vitamine sowie RNA, DNA und Ribosomen umfasst. Der enzymatische Komplex der Mitochondrien beschleunigt den komplexen und miteinander verbundenen Mechanismus biochemischer Reaktionen, die zur Bildung von ATP führen. In diesen Organellen werden Zellen mit Energie versorgt – die Energie chemischer Nährstoffbindungen wird im Prozess der Zellatmung in hochenergetische ATP-Bindungen umgewandelt. In den Mitochondrien erfolgt der enzymatische Abbau von Kohlenhydraten, Fettsäuren und Aminosäuren unter Freisetzung von Energie und deren anschließender Umwandlung in ATP-Energie. Die angesammelte Energie wird für Wachstumsprozesse, für neue Synthesen etc. aufgewendet. Mitochondrien vermehren sich durch Teilung und leben etwa 10 Tage, danach werden sie zerstört.
Endoplasmatisches Retikulum
Das endoplasmatische Retikulum ist ein Netzwerk aus Kanälen, Röhren, Vesikeln und Zisternen, die sich im Zytoplasma befinden. Es wurde 1945 vom englischen Wissenschaftler K. Porter entdeckt und ist ein Membransystem mit ultramikroskopischer Struktur.
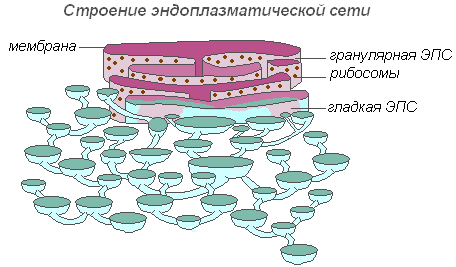
Struktur des endoplasmatischen Retikulums
Das gesamte Netzwerk ist mit der äußeren Zellmembran der Kernhülle zu einem Ganzen verbunden. Es gibt glatte und raue ER, die Ribosomen tragen. Auf den Membranen des glatten ER befinden sich Enzymsysteme, die am Fett- und Kohlenhydratstoffwechsel beteiligt sind. Dieser Membrantyp überwiegt in Samenzellen, die reich an Speichersubstanzen (Proteine, Kohlenhydrate, Öle) sind; an der körnigen EPS-Membran sind Ribosomen befestigt, und bei der Synthese eines Proteinmoleküls wird die Polypeptidkette mit Ribosomen in den EPS-Kanal eingetaucht. Die Funktionen des endoplasmatischen Retikulums sind sehr vielfältig: Stofftransport sowohl innerhalb der Zelle als auch zwischen benachbarten Zellen; Aufteilung einer Zelle in separate Abschnitte, in denen verschiedene physiologische Prozesse und chemische Reaktionen gleichzeitig ablaufen.
Ribosomen
Ribosomen sind zelluläre Organellen ohne Membran. Jedes Ribosom besteht aus zwei Partikeln, deren Größe nicht identisch ist und die in zwei Fragmente geteilt werden können, die nach der Vereinigung zu einem ganzen Ribosom weiterhin die Fähigkeit zur Proteinsynthese behalten.
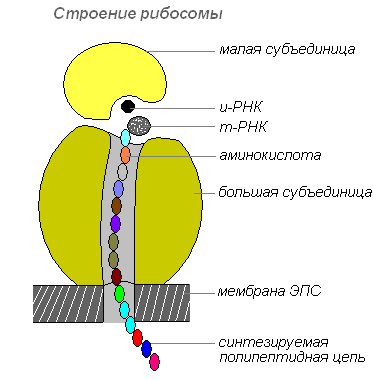
Ribosomenstruktur
Ribosomen werden im Zellkern synthetisiert, verlassen ihn dann und wandern in das Zytoplasma, wo sie an der Außenfläche der Membranen des endoplasmatischen Retikulums befestigt werden oder sich frei befinden. Abhängig von der Art des zu synthetisierenden Proteins können Ribosomen einzeln funktionieren oder zu Komplexen – Polyribosomen – zusammengefasst sein.
Die Zellwand ist ein Derivat des Protoplasten, d.h. entsteht während seiner Lebensaktivität (Abb. 61). Es verleiht der Zelle eine bestimmte Form, schützt den Protoplasten und verhindert durch Widerstand gegen den intrazellulären Druck den Zellbruch. Zellwände erfüllen die Funktionen des inneren Skeletts einer Pflanze und verleihen ihren Organen die notwendige mechanische Festigkeit.
Zellwände lassen Sonnenlicht gut durch und Wasser und darin gelöste Mineralien bewegen sich leicht durch sie hindurch. Zwischen den Wänden benachbarter Zellen gibt es Mittelplatte - Pektinschicht, die eigentlich eine interzelluläre Substanz ist und die Wände benachbarter Zellen zusammenhält. An den Stellen, an denen sich die Zellwände benachbarter Zellen nicht schließen, sind sie mit Wasser gefüllt Interzellularräume. Als Prozess der Zerstörung der Interzellularsubstanz wird bezeichnet, wodurch die Wände benachbarter Zellen getrennt werden Mazeration. Es ist eine natürliche Mazeration zu beobachten
Reis. 61.
A- Diagramm der Struktur der Zellwand; B- Diagramm der Beteiligung des Golgi-Apparats am Aufbau der Zellwand; IN- detaillierter Aufbau der Zellwand: 1 - Mittelplatte; 2 - es ist Zeit; 3 - Sekundärwand;
- 4 - Primärwand; 5 - Diktyosom; 6 - Golgi-Vesikel;
- 7-Plasmalemma; 8- Zellenwand; 9- Makrofibrille;
- 10-Mikrofibrille; 11 - Mizelle; 12 - Zellulosemolekül;
- 13 – Struktur eines Cellulosemolekülfragments
in überreifen Früchten von Äpfeln, Ebereschen, Melonen usw. Eine künstliche Mazeration wird beispielsweise beim Einweichen von Flachsstielen durchgeführt, um Bastfasern von ihnen zu trennen; Es tritt auch bei der Wärmebehandlung von Früchten auf.
Die Zellwand enthält Polysaccharide: Pektine, Hemizellulose und Zellulose. Sehr lange Cellulosemoleküle sind geordnet parallel zueinander angeordnet (jeweils 40–60) und bilden sich Mizellen. Mizellen werden in Bündeln gesammelt - Mikrofibrillen, stellt die Grundstruktureinheit der Cellulose dar. Mikrofibrillen wiederum werden zu kombiniert Makrofibrillen- sehr dünne Fasern ungewisser Länge. Zellulosemakrofibrillen werden in stark gewässertes Wasser eingetaucht Matrix, bestehend aus Pektinen, Hemizellulosen und einigen anderen Stoffen. Die Festigkeit der Zellwand wird durch elastische Zellulose-Mikrofibrillen gegeben, deren Zugfestigkeit der von Stahl nahe kommt. Die Stärke und Elastizität der Zellwand liegt ihrer Fähigkeit zugrunde, sich reversibel zu dehnen. Dank Pektinen und Hemizellulose ist die Zellwand sehr wasserdurchlässig – Wasser und darin gelöste Stoffe wandern problemlos von Zelle zu Zelle.
Die Zellwand grenzt außen an das Plasmalemma an, das aktiv an seinem Wachstum teilnimmt. Moleküle aus Pektinen, Hemizellulose, Zellulose und anderen Substanzen werden synthetisiert und in den Dictyosomen-Zisternen des Golgi-Apparats angesammelt. Golgi-Vesikel transportieren sie zur Peripherie des Protoplasten – zum Plasmalemma. An der Kontaktstelle zwischen Vesikel und Plasmalemma löst sich dieses auf und der Inhalt der Vesikel, sobald er sich außerhalb des Plasmalemmas befindet, wird zum Aufbau der Zellwand verwendet. Die Vesikelmembran stellt nicht nur die Integrität des Plasmalemmas wieder her, sondern sorgt auch für dessen Oberflächenwachstum. Das Wachstum der Zellwand erfolgt aufgrund der enzymatischen Aktivität des Plasmalemmas.
Als Wände werden die sich teilenden und wachsenden Zellen bezeichnet primär. Sie enthalten viel Wasser (60-90 %), ihre Trockenmasse wird von Pektinen und Hemizellulose dominiert – der Zelluloseanteil beträgt nicht mehr als 30 %. Wenn sich eine Zelle in der Telophase der Mitose teilt, wird die Mutterzelle durch die Bildung eines Septums in ihrer Äquatorialebene in zwei Tochterzellen geteilt – Mittelplatte. Auf beiden Seiten der Mittelschicht beginnt jede der beiden Tochterzellen, ihre eigene primäre Zellwand zu bilden. Das Wachstum der Mittelplatte und der Primärwände der beiden Tochterzellen erfolgt in zentrifugaler Richtung – vom Zentrum der Mutterzelle bis zu ihrer Peripherie. Die Mittelplatte ist sehr dünn und besteht aus Pektin.
Die durch die Teilung entstehende neue Zelle beginnt zu wachsen und ihr Volumen kann sich um das Hundertfache oder mehr vergrößern. Das Zellwachstum erfolgt hauptsächlich durch Verstauchungen aufgrund der Aufnahme von Wasser und der Vergrößerung des Vakuolenvolumens. Durch den entstehenden Innendruck dehnt sich die Primärwand, in die Cellulosemicellen, Pektine und Hemicellulose leicht eingebettet werden. Methode des Zellwandwachstums nach Implementierung Baumaterial zwischen bestehenden Bauwerken genannt Invagination.
In der primären Zellwand gibt es zunächst dünnere Bereiche, in denen die Zellulosefibrillen lockerer angeordnet sind – primäre Porenfelder. Die primären Porenfelder der Wände zweier benachbarter Zellen fallen normalerweise zusammen. Hier wandern die Tubuli des endoplasmatischen Retikulums von einer Zelle zur anderen – Plasmodesmen. Die Wege, auf denen Plasmodesmen von einer Zelle zur anderen gelangen, werden genannt plasmodesminale Tubuli. Durch diese Tubuli sind die Hyaloplasmen benachbarter Zellen miteinander verbunden. Plasmodesmen übernehmen den interzellulären Transport von Stoffen (Hormone, Aminosäuren, ATP, Zucker usw.). Die mit Hilfe von Plasmodesmen zu einem Ganzen vereinten Protoplasten von Körperzellen werden Symplasten genannt. Als Transport von Stoffen entlang von Plasmodesmen wird bezeichnet symplastisch.(Man nennt die Gesamtheit der Zellwände, der Mittelschicht und der Interzellularräume Apoplast, geht auf ihnen apoplastisch Transport von Stoffen.)
Nachdem das Zellwachstum abgeschlossen ist, kann seine Primärwand dünn bleiben (bei Bildungsgewebezellen) oder beginnen, dicker zu werden (bei permanenten Gewebezellen). Das Wachstum der Zellwanddicke nennt man sekundäre Verdickung. Dadurch lagert es sich an der Innenfläche der Primärwand ab. Sekundärwand was wächst Apposition- Aufbringen von Zellulosemizellen auf eine bestehende Wand. In diesem Fall befinden sich die jüngsten Schichten der sekundären Zellwand neben dem Plasmalemma. Die sekundäre Zellwand übernimmt vor allem tragende und mechanische Funktionen. Seine Zusammensetzung enthält deutlich weniger Wasser als die primäre und in der Trockenmasse überwiegt Zellulose (bis zu 50 %). Beispielsweise kann in den Sekundärwänden einzelliger Haare aus Baumwoll- und Flachsbastfasern der Zellulosegehalt bis zu 95 % betragen.
Die sekundäre Verdickung der Zellwand erfolgt ungleichmäßig. Abschnitte der sekundären Zellwand an den Stellen der primären Porenfelder bleiben in der Regel unverdickt. Solche nicht verdickten Bereiche der Zellwand nennt man manchmal. Poren in den Wänden zweier benachbarter Zellen fallen in der Regel zusammen und bilden sich ein paar Mal. Der durch ein Porenpaar gebildete Porenkanal ist verstopft Verschlussfilm der Pore - ein Septum, bestehend aus einer mittleren Lamina und zwei Primärwänden benachbarter Zellen. Der Verschlussfilm der Pore wird von zahlreichen Plasmodesmen durchdrungen, durch die Plasmodesmen hindurchtreten.
Poren werden unterschieden einfach Und begrenzt(Abb. 62). Bei einfachen Poren ist der Durchmesser ihres Porenkanalabschnitts über die gesamte Länge gleich, d. h. es hat eine zylindrische Form. Einfache Poren sind typisch für Parenchymzellen. Umrandete Poren sind charakteristisch für die Wände von Zellen, die Wasser mit gelösten Mineralien leiten – Tracheiden und Gefäßsegmente. Bei solchen Poren hat ihr Teil des Porenkanals die Form eines Trichters, dessen breite Seite an den Verschlussfilm der Pore angrenzt.
In den Zellen leitender Gewebe von Nadelbäumen ist der Porenverschlussfilm nur an den Rändern wasserdurchlässig, da sein zentraler scheibenförmiger Teil verdickt und verholzt ist Torus - lässt kein Wasser durch. Der Torus übernimmt die Rolle einer Klappe. Ist der Wasserdruck in benachbarten Zellen nicht gleich, wird der Schließfilm abgelenkt und der Torus blockiert die Bewegung des Wassers durch den Porenkanal.

Reis. 62.
A- einfach; B- begrenzt; IN- halbkantig:
1 - Abschlussfilm; 2 - Porenkanal; 3 - Torus
Zusätzlich zu den Poren befinden sich in den Wänden wasserleitender Zellen Zähnung- Durchgangslöcher (Gefäßsegmente, wasserspeichernde Zellen von Torfmoos).
Veränderungen der Zellwand. Abhängig von den Funktionen der Zelle kann ihre Wand durch die Ablagerung von Substanzen in ihr verändert werden. Seine üblichen Modifikationen sind: Verholzung, Suberisierung, Kutinisierung, Mineralisierung und Schleimbildung.
Zellwandverholzung, oder Verholzung, entsteht durch die Ablagerung von Lignin, einem aromatischen Stoff mit komplexer chemischer Struktur, in den intermicellaren Räumen. Die Festigkeit und Härte der Wand nehmen zu, ihre Elastizität nimmt jedoch ab. Verholzte Wände sind in der Lage, Wasser und Luft durchzulassen. Bei einer verholzten Zellwand bleibt der Protoplast der Zelle möglicherweise am Leben, stirbt jedoch normalerweise ab. Bei manchen Gehölzen reichert sich bis zu 30 % Lignin im Holz an. Lignin kann sich auch in den Zellwänden alternder Grastriebe ansammeln, was deren Futterwert deutlich verringert und den Zeitpunkt der Heuernte bestimmt. Bei der Gewinnung von Zellulosezellstoff aus Holz, das für die Papierherstellung notwendig ist, wird eine künstliche Entholzung durchgeführt. Eine natürliche Verholzung der Zellwand ist möglich, aber selten.
Probenahme, oder Suberinisierung, - Ablagerung einer persistenten, fettähnlichen, amorphen Substanz Suberin (hydrophobes Polymer) in der Zellwand. Suberisierte Zellwände sind für Gase und Wasser undurchlässig, was zum Absterben des Protoplasten führt. Zellen mit suberisierten Wänden schützen Pflanzen zuverlässig vor Wasserverlust, extremen Temperaturen, pathogenen Bakterien und Pilzen.
Kutinisierung - Ablagerung von Cutin (einer Substanz mit ähnlicher chemischer Zusammensetzung wie Suberin) in den Zellwänden. Cutin wird normalerweise in den oberflächlichen Schichten der Außenwände von Zellen und auf deren Oberflächen abgelagert. In Form eines Films – der Kutikula – bedeckt es beispielsweise die Oberfläche der Zellen des Hautgewebes – der Epidermis.
Mineralisierung Die Zellwand wird durch die Ablagerung von Kalzium- und Kieselsäuresalzen zerstört. Diese Stoffe verleihen der Wand Härte und Sprödigkeit. Der Mineralisierungsprozess kommt besonders gut in den Zellwänden der Epidermis von Getreide-, Seggen- und Schachtelhalmsprossen zum Ausdruck. Aus diesem Grund empfiehlt es sich, die Triebe von Seggen und Gräsern vor der Blüte zu mähen – später werden sie durch die starke Mineralisierung gröber, was die Qualität des Heus beeinträchtigt.
Schleim- Umwandlung von Zellulose und Zellwandpektinen in spezielle Polysaccharide - Schleim und Zahnfleisch, die bei Kontakt mit Wasser stark aufquellen können. Ein Abrutschen der Wand wird bei Samenschalenzellen beobachtet, beispielsweise bei Quitte, Flachs, Gurke und Wegerich. Klebriger Schleim kann die Samenverbreitung fördern (Wegerich); Wenn die Samen keimen, absorbieren und speichern die Schleimstoffe Wasser und schützen sie so vor dem Austrocknen. In der Wurzelkappe fungiert Schleim als Gleitmittel und erleichtert den Durchgang der Wurzel zwischen Erdklumpen. Schleim und Zahnfleisch können in erheblichen Mengen entstehen, wenn sich Zellwände aufgrund ihrer Schädigung auflösen. Kirschen und Pflaumen bilden oft Gummi, wenn Zweige und Stämme verletzt werden. Der sogenannte Kirschkleber ist ein in Form von Perlen aushärtender Kaugummi, der die Oberfläche von Wunden und Frostlöchern bedeckt und das Eindringen von Infektionen verhindert. Schleim dieser Art nennt man Gumose und gilt als pathologisches Phänomen.
Da sekundäre Zellwände als inneres Skelett einer Pflanze fungieren und ihren Organen die nötige Festigkeit verleihen (was besonders wichtig für Landpflanzen ist), können sie sich häufig – lokal oder vollständig – erheblich verdicken, um ihnen eine größere Festigkeit zu verleihen Gewebe und damit zum Pflanzenorgan. Durch die Ablagerung von Zellulose kommt es zu einer Verdickung der Zellwand.
Die Funktionen von Zellen werden oft ausschließlich von ihren Wänden übernommen, da die Protoplasten der Zellen absterben. Dies betrifft die Korkzellen,

Reis. 63.
Tracheiden, Gefäßsegmente, mechanische Gewebefasern. Das Holz, das die meisten riesigen Baumstämme einnimmt, besteht beispielsweise überwiegend aus verholzten Zellwänden, deren Protoplasten längst abgestorben sind.
Zellwände spielen in unserem Leben eine große Rolle. Aus ihnen werden textile Rohstoffe (Baumwollsamenhaare, Flachsfasern etc.) und Rohstoffe für die Herstellung von Seilen und Schnüren (Hanffasern, Seilfasern, Sisal etc.) gewonnen. Aus Zellwänden gewonnene Zellulose wird zur Herstellung von Papier (Fichten- und Espenholz), Acetatseide, Viskose, Kunststoffen, Zellophan und vielem mehr verwendet. Kork ist ein Gewebe aus abgestorbenen Zellen mit suberisierten Wänden und wird seit langem als wertvolles wasser- und luftdichtes Wärmedämmmaterial verwendet und wird zunehmend im modernen Bauwesen eingesetzt.
 (1 Bewertungen im Durchschnitt: 5,00 von 5)
(1 Bewertungen im Durchschnitt: 5,00 von 5) 

